
Rekayasa Mikrojaringan Glomerulus Ginjal 3D untuk Memodekan Penyakit yang Berpusat pada Podosit guna Validasi Target Obat Baru
Abstrak
Podositopati adalah kelompok penyakit ginjal yang beragam yang ditandai dengan cedera podosit, yang menyebabkan proteinuria dan penurunan fungsi ginjal. Cedera ini sering mengganggu dinamika sitoskeletal dan adhesi seluler, yang menyebabkan disfungsi glomerulus. Model in vitro saat ini gagal meniru secara akurat organisasi tiga dimensi (3D) dan mekanika jaringan ginjal, yang menghambat pemahaman patofisiologi podosit dan pengembangan terapi. Dalam penelitian ini, mikrotisis 3D dikembangkan yang mereplikasi struktur dan mekanika penghalang filtrasi glomerulus, yang memungkinkan pemodelan cedera podosit yang diinduksi secara kimia dan genetika untuk validasi target obat. Mikrotisis ini mensimulasikan struktur tiga lapis glomerulus dan peregangan mekanis hemodinamik, yang menyediakan platform untuk mengevaluasi jalur pensinyalan mekanobiologis dan dinamika podosit yang relevan. Kekuatan seluler kolektif diukur untuk menilai ketahanan podosit terhadap cedera genetik atau kimia. Sebagai bukti konsep, cedera podosit dimodelkan melalui ekspresi berlebih transient receptor potantial canonical 6 (TRPC6), target yang divalidasi dalam podositopati, dan dievaluasi oleh inhibitor TRPC6 SAR7334. Hasilnya menunjukkan hilangnya kekuatan kontraksi podosit setelah ekspresi berlebih TRPC6, dengan pemulihan setelah pengobatan. Hal ini menyoroti potensi jaringan mikro glomerulus untuk memodelkan mekanopatofisiologi podosit dan berfungsi sebagai platform yang kuat untuk menyaring terapi baru.
1 Pendahuluan
Glomerulus manusia adalah antarmuka yang sangat terspesialisasi yang berfungsi sebagai unit penyaringan ginjal. Glomerulus dipertahankan melalui interaksi di antara komponen-komponen utamanya: sel endotel glomerulus (GEC), membran dasar glomerulus (GBM), dan podosit. [ 1 ] Podosit adalah sel epitel yang berdiferensiasi secara terminal yang membungkus kapiler glomerulus. Dengan prosesus kaki yang saling bertautan (FP), podosit membentuk tulang punggung arsitektur penghalang penyaringan glomerulus (GFB) untuk memberikan ultrafiltrasi dan stabilitas mekanis pada jaringan di bawahnya.
Dalam struktur tiga lapis ini, GEC dan podosit terletak pada GBM yang disekresikan sendiri yang terdiri dari komponen matriks ekstraseluler (ECM) yang unik: kolagen IV (Col-IV) dan laminin, dengan isoformnya berubah saat GBM matang. [ 2 , 3 ] Kekuatan utama yang memengaruhi podosit meliputi tegangan geser cairan transmural dari ultrafiltrat dan tegangan tarik melingkar dari peregangan hemodinamik. [ 4 ] Untuk mempertahankan filtrasi glomerulus, podosit harus menahan dan beradaptasi dengan kekuatan mekanis ini dengan merombak sitoskeletonnya dan memperkuat titik adhesi sel-matriks. Gagal melakukannya biasanya dikaitkan dengan keadaan penyakit dan penurunan fungsi ginjal. [ 5 ]
Podositopati merupakan kelas penyakit ginjal heterogen di mana cedera podosit menyebabkan proteinuria dan penurunan fungsi ginjal. Pada kelas penyakit ini, cedera podosit dapat berasal dari kondisi genetik, imun, infeksi, toksik, metabolik, atau hemodinamik dan mengakibatkan terganggunya dinamika aktin sitoskeleton dan penghapusan FP. [ 6 ] Contoh podositopati genetik adalah mutasi perolehan fungsi pada saluran TRPC6, yang dilaporkan menyebabkan kerusakan podosit melalui kelebihan kalsium intraseluler, yang menyebabkan penghapusan FP dan pelepasan podosit dari GBM, yang menyebabkan proteinuria dan glomerulosklerosis. [ 7 , 8 ]
Mengembangkan model in vitro yang merangkum arsitektur barier filtrasi glomerulus manusia dan mekanobiologi podosit akan sangat memudahkan pemahaman mekanistik tentang podositopati dan memungkinkan pengembangan terapi yang ditargetkan pada podosit. Hingga saat ini, model sferoid kultur bersama 3D dari glomerulus ginjal, yang dikenal sebagai GlomSpheres, telah dikembangkan untuk meniru struktur fisik dan molekuler glomerulus yang spesifik, yang memungkinkan penggunaannya dalam skrining obat dengan hasil tinggi. [ 9 ] Selain itu, organoid ginjal yang berasal dari sel punca pluripoten yang diinduksi berhasil mereplikasi struktur utama seperti nefron termasuk podosit (saling berhubungan erat) dan sel mesangial; namun, mereka tidak menunjukkan GEC, GBM dewasa, atau isyarat mekanis yang menyerupai kondisi hemodinamik. [ 10 ] Sistem mikrofluidik, tempat podosit dan GEC terletak pada sisi berlawanan dari membran berpori, menggabungkan gaya tegangan geser media dan, dalam kasus tertentu, peregangan podosit, yang memungkinkan sekresi GBM yang lebih matang. Podosit tetap datar dalam sistem ini dan tidak menciptakan tulang punggung arsitektur melengkung yang mengelilingi endotelium. Selain itu, membran fisik antara sel endotel dan podosit dapat membatasi interaksi mereka. [ 11 ] Baru-baru ini, model mikrofluidik yang menampilkan deposisi GBM yang dapat disesuaikan dan kultur bersama 3D podosit dengan GEC menunjukkan bahwa kontrol yang tepat atas ketebalan GBM dapat secara efektif meniru filtrasi fisiologis dan respons proteinurik patofisiologis dari GFB. Selain itu, kedekatan GEC dan podosit dalam model ini meningkatkan komunikasi antar sel yang dinamis. [ 12 ] Namun, model ini masih kekurangan komponen mekanis penting dari GFB.
Di sini, kami memperkenalkan model seluler 3D tingkat lanjut, yang mereplikasi arsitektur GBM tiga lapis, termasuk pembungkus podosit di sekitar endotelium, dengan pengganti mekanis terintegrasi untuk gaya hemodinamik guna mensimulasikan mekanofisiologi dan patofisiologi podosit yang terkait dengan podositopati. Dengan integrasi titik akhir yang relevan dengan penyakit (yaitu, gaya kontraktil podosit dan pelepasan podosit dari GBM), platform ini dapat digunakan untuk meningkatkan pemahaman mekanistik kita tentang cedera podosit dan berfungsi sebagai platform baru untuk memvalidasi target podosit dan menguji senyawa. Strategi dua langkah kami memungkinkan pembentukan jaringan mikro endotel 3D dalam gel kolagen-I diikuti oleh pembungkus eksternal jaringan mikro oleh podosit. Dalam konfigurasi ini, sel-sel merombak lingkungannya dengan mengeluarkan komponen GBM termasuk Col-IV dan laminin. Dalam model ini, isyarat mekanis juga beraksi, karena jaringan mikro yang terbentuk di sekitar pilar mikro yang fleksibel berada di bawah tekanan karena resistensi pilar terhadap tarikan oleh sel-sel. Ketegangan statis juga dapat dikombinasikan dengan peregangan siklik sebagai pengganti modulasi tekanan hemodinamik dengan tujuan merangkum semua tegangan tarik melingkar yang dirasakan oleh podosit selama kondisi normal dan hipertensi. Pemantauan pengukuran gaya dan cakupan podosit pada mikrotisu, kami dapat menilai ketahanan biomekanik podosit setelah cedera dan pengobatan dengan senyawa. Untuk mengonfirmasi relevansi model kami, kami menguji podosit yang diabadikan dengan ekspresi berlebih TRPC6 yang dapat diinduksi doksisiklin (Dox) untuk mereplikasi mutasi perolehan fungsi TRPC6, yang mengakibatkan kerusakan sitoskeletal podosit dan hilangnya gaya seluler. Kami mengonfirmasi dampak negatif dari ekspresi berlebih TRPC6 pada kesehatan podosit dan menunjukkan peran protektif gaya seluler saat mikrotisu diobati dengan penghambat TRPC6 SAR7334. Ini memvalidasi model kami sebagai platform yang berharga untuk validasi target dan pengujian senyawa.
2 Hasil dan Pembahasan
2.1 Podosit Membungkus Matriks Kolagen yang Dirakit Sendiri, Mengandung Sel Endotel, Menyebabkan Pembentukan Mikrojaringan Glomerulus 3D
Penghalang filtrasi glomerulus adalah struktur tiga lapis yang dibentuk oleh sel-sel endotel berfenestrasi, membran dasar glomerulus, dan podosit yang saling bertautan. [ 1 ] Podosit memberikan stabilitas mekanis glomerulus dengan melekat pada GBM dan membungkus erat mikrokapiler glomerulus ( Gambar 1 A ). Untuk merangkum struktur 3D ini, kami menggunakan perangkat polidimetilsiloksan (PDMS) yang lentur yang berisi serangkaian sumur mikro dengan dimensi 3,6×1,8×2 mm (pxlxt) masing-masing dengan dua mikropilar fleksibel vertikal (diameter 500 µm dengan jarak pusat-ke-pusat 1,8 mm) (Gambar S1A , Informasi Pendukung). [ 13 ] Ketika campuran sel endotel manusia dengan protein matriks ekstraseluler, kolagen-I dalam kasus ini, ditempatkan dalam sumur mikro individual, sel-sel mulai menyebar dan secara kolektif memberikan gaya traksi di dalam gel terpolimerisasi. Kekuatan traksi oleh aktin cytoskeleton kemudian mengarah pada pemadatan ECM yang mengakibatkan pembentukan jaringan mikro endotel yang tersuspensi di antara pilar setelah 24 jam (Gambar 1B ). Karena pilar mikro fleksibel, maka gaya kontraktil yang dihasilkan jaringan dapat dihitung dengan mengukur defleksi pilar (Gambar 1C ). Dengan demikian, model ini memungkinkan kita untuk memantau perubahan gaya seluler sebagai indikasi reorganisasi dan kerusakan cytoskeletal. Selain itu, resistensi pilar mikro terhadap pemadatan dan gaya yang dihasilkan oleh sel memastikan ketegangan konstan yang diterapkan pada jaringan mikro selama periode kultur.
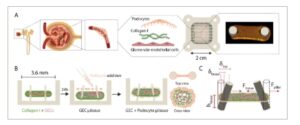
Gambar 1
Buka di penampil gambar
Kekuatan Gambar
Representasi skematis dari strategi pengembangan jaringan mikro glomerulus. A) Penggambaran struktur nefron, glomerulus, dan GFB. GFB terdiri dari GEC berfenestrasi, GBM, dan podosit. Jaringan mikro yang meniru GFB dibentuk dalam perangkat PDMS. B) Metodologi untuk menghasilkan jaringan mikro. GEC yang tertanam dalam kolagen-I memungkinkan pembentukan awal jaringan mikro GEC, yang selanjutnya dimatangkan melalui penggabungan podosit, yang menyediakan cakupan dan pembangkitan gaya utama. C) Ilustrasi jaringan yang menghasilkan gaya dan membelokkan pilar. Dibuat dalam BioRender. Yasa, I. (2025) https://BioRender.com/brlp530 .
Kepadatan sel memainkan peran penting dalam pembentukan mikrotisu dengan memadatkan matriks di sekitar mikropilar. [ 14 ] Kami menguji kepadatan sel endotel yang bervariasi dari 1,5× 104 , 2× 104 , 3×104 per mikrotisu dan mengamati bahwa mikrotisu dipadatkan lebih cepat, dalam waktu 24 jam, ketika kepadatan sel adalah 3×104 baik dengan sel endotel vena umbilikalis manusia (HUVEC) dan GEC (Gambar S1B , Informasi Pendukung). Kepadatan ini adalah minimum yang diperlukan untuk pembentukan mikrotisu. Meskipun kepadatan sel yang lebih tinggi berpotensi mempercepat proses, dalam kultur yang diperpanjang, hal itu dapat meningkatkan risiko pelepasan dini mikrotisu dari mikropilar karena kekuatan yang lebih tinggi yang terlibat. Oleh karena itu, kami melanjutkan dengan kepadatan sel minimum yang memungkinkan pembentukan mikrotisu endotel dalam waktu 24 jam. Bahasa Indonesia: Setelah jaringan mikro endotel terbentuk, kami memperkenalkan podosit yang diabadikan secara bersyarat manusia (ciPods) ke dalam sistem dan menyelidiki bagaimana, dengan membungkus permukaan jaringan mikro, mereka menghasilkan jaringan mikro glomerulus. Podosit diberi label dengan Calcein-Green untuk memvisualisasikan adhesi dan penyebarannya secara spasial. Kami mengamati perlekatan podosit ke permukaan jaringan mikro endotel ( Gambar 2A ) dan morfologi jaringan mikro endotel dan glomerulus sangat dapat direproduksi (Gambar S2 , Informasi Pendukung). Menariknya, perlekatan podosit sebagian besar diamati dengan jaringan mikro endotel, karena tidak ada adhesi yang terdeteksi dengan jaringan mikro fibroblas yang dibentuk oleh fibroblas dermal manusia (Gambar S3 , Informasi Pendukung). Namun, penyelidikan lebih lanjut diperlukan untuk sepenuhnya memahami perilaku ini dan mengonfirmasi spesifisitas cakupan podosit. Kemudian, kami mengukur perubahan dalam lebar jaringan mikro dan menemukan bahwa podosit lebih lanjut memadatkan jaringan mikro sebesar 26 ± 4% dengan membungkus dibandingkan dengan yang hanya endotel (Gambar S4 , Informasi Pendukung). Perbedaan morfologi ini dikaitkan dengan peningkatan yang signifikan dalam gaya kontraktil (Gambar 2B ). Oleh karena itu, kami mengukur gaya statis yang diberikan oleh sel untuk memadatkan jaringan mikro baik dalam jaringan mikro endotel maupun dalam jaringan mikro glomerulus. Kami menemukan generasi gaya yang secara signifikan lebih tinggi dalam jaringan mikro glomerulus dibandingkan dengan yang endotel, termasuk jaringan mikro HUVEC dan GEC (Gambar 2B ). Setelah mengonfirmasi pembentukan jaringan mikro tanpa perbedaan yang nyata oleh berbagai sumber sel endotel, kami melanjutkan hanya dengan GEC sebagai sel yang paling relevan dalam kapiler glomerulus.
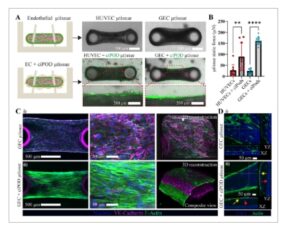
Gambar 2
Buka di penampil gambar
Kekuatan Gambar
Mikrotisu glomerulus yang direkayasa. A) Citra brightfield dari mikrotisu yang dihasilkan oleh berbagai sumber sel endotel, HUVEC, dan GEC. CiPOD berlabel Calcein Green divisualisasikan menempel pada permukaan mikrotisu. B) Pengukuran gaya mikrotisu dilakukan pada hari ke-4 kultur, 3 hari setelah penambahan ciPOD. Mikrotisu glomerulus menunjukkan gaya yang jauh lebih tinggi dibandingkan dengan mikrotisu endotel. Data dilaporkan sebagai mean ± SD, n ≥ 9 mikrotisu dan ** P < 0,01, **** P < 0,0001 jika dibandingkan dengan ANOVA satu arah dengan uji perbandingan berganda Tukey. C) Citra konfokal fluoresen terproyeksi dua dimensi (2D) representatif dan terekonstruksi 3D dari mikrotisu GEC (i) dan glomerulus (GEC + ciPOD) (ii), masing-masing. GEC dibedakan berdasarkan ekspresi VE-Cadherin dan ciPOD berdasarkan serat aktinnya. D) Pencitraan harmonik kedua kolagen-I di kedua mikrotisu GEC (i) dan mikrotisu glomerulus (ii), yang menunjukkan organisasi matriks dan penghalang antara podosit (panah kuning) dan GEC (panah merah) di mikrotisu glomerulus (ii).
Meskipun gaya yang lebih tinggi dalam jaringan mikro glomerulus menunjukkan kontribusi yang jelas dari penambahan podosit dan pembungkusan jaringan mikro berikutnya, kami mempertanyakan apakah peningkatan pembentukan gaya ini disebabkan oleh faktor parakrin yang disekresikan oleh podosit yang dapat memengaruhi perilaku dan kontraktilitas sel endotel. Untuk membedakan kontribusi podosit terhadap gaya yang diukur, kami menguji efek pengobatan trombin dan faktor pertumbuhan endotel vaskular-A (VEGF-A) karena keduanya diketahui menginduksi gaya traksi dan kontraktilitas sel endotel. [ 15 , 16 ] Kami mengobati jaringan mikro GEC dengan 10 U ml −1 trombin selama 40 menit dan mengukur gaya setiap 5 hingga 15 menit dan membandingkan hasilnya dengan jaringan mikro glomerulus yang dibungkus oleh podosit. Kami mengamati peningkatan yang signifikan dalam pembentukan gaya, ca. 4 lipatan dengan 67 ± 14 µN, ketika jaringan mikro GEC diobati dengan trombin setelah 5 menit. Gaya yang dihasilkan tidak menunjukkan perubahan signifikan selama 40 menit dan tetap ≈73 ± 8 µN (Gambar S5A , Informasi Pendukung). Ketika kelompok yang diobati dengan trombin dibandingkan dengan jaringan mikro glomerulus, kami menemukan perbedaan signifikan dengan jaringan mikro glomerulus yang mengerahkan gaya 160 ± 16 µN. Sebaliknya dengan efek trombin, ketika jaringan mikro GEC diobati dengan 100 ng ml −1 VEGF-A, perubahan gaya terasa lebih kecil, dan tidak ditemukan signifikansi setelah 24 jam dan 48 jam antara jaringan mikro GEC yang tidak diobati dan diobati (Gambar S5B , Informasi Pendukung). Hasil-hasil ini mengonfirmasi pemadatan lebih lanjut jaringan mikro glomerulus oleh podosit karena kekuatan yang jauh lebih tinggi dihitung dengan adanya podosit dibandingkan dengan perawatan VEGF-A dan trombin.
Kami kemudian menyelidiki secara dekat komponen-komponen dan organisasi spasial dari mikrotissu dengan imunofluoresensi. Kami mewarnai mikrotissu untuk VE-Cadherin, sebagai penanda sel endotel, dan F-aktin untuk membedakan podosit. Kami mengamati adhesi sel-ke-sel GEC yang membentuk lapisan endotelium kompak dengan organisasi aktin yang kurang padat (Gambar 2C-i ). Di sisi lain, dalam literatur, podosit terbukti memberikan integritas mekanis dan arsitektur melalui pameran sitoskeleton unik yang dicirikan oleh polaritas sel dan jaringan tonjolan yang luas. [ 17 ] Kami juga selanjutnya mengonfirmasi pembungkusan mikrotissu oleh podosit dengan sitoskeleton aktin yang sangat terorganisir yang juga selaras dengan gaya yang dihasilkan (Gambar 2C-ii ; Gambar S4C , Informasi Pendukung). Intensitas dan perbedaan jaringan dalam serat stres terbukti dari gambar confocal jika dibandingkan dengan mikrotissu GEC. Selain itu, keberadaan lapisan GEC di bawah podosit di sekitarnya dikonfirmasi oleh gambar confocal rekonstruksi 3D dari jaringan mikro glomerulus (Gambar 2C-ii ). Analisis mikroskopi dua-foton memberikan informasi lebih lanjut tentang organisasi spasial jaringan mikro glomerulus. Kami mencitrakan kolagen-I dengan generasi harmonik kedua dan memvisualisasikan bahwa kolagen-I terletak di antara lapisan GEC dan podosit, dengan cara yang mirip dengan membran dasar glomerulus. Dengan tidak adanya podosit, GEC malah bermigrasi dan menata ulang untuk membentuk jaringan paling atas (Gambar 2D ; Gambar S6 , Informasi Pendukung), menyusun diri mereka sendiri menjadi lapisan tunggal kompak eksternal dengan sambungan sel-ke-sel yang utuh, dan jaringan sel internal yang kurang terorganisir yang tersebar dalam ECM. Secara keseluruhan, hasil-hasil ini menunjukkan bahwa strategi kami memungkinkan pembentukan jaringan mikro yang sangat menyerupai antarmuka jaringan-jaringan normal dari penghalang filtrasi glomerulus dalam 3D.
2.2 Peningkatan Ekspresi dan Sintesis Komponen GBM dalam Mikrotisu 3D
Untuk memahami dampak organisasi 3D podosit dengan crosstalk endotel pada ekspresi gen, pertama-tama kami menguji tingkat ekspresi podocalyxin (PODXL) dan synaptopodin (SYNPD). PODXL adalah protein permukaan sel glikosilasi yang diekspresikan dalam podosit dan GEC dan diperlukan untuk pengembangan dan pemeliharaan FP. SYNPD adalah protein terkait aktin dalam podosit yang memainkan peran penting dalam pengembangan dan pemeliharaan aparatus kontraktil podosit. Ketika kami membandingkan jaringan mikro glomerulus dengan podosit berlapis 2D, kami mendeteksi peningkatan ekspresi PODXL dan SYNPD dalam waktu 3 hari setelah menambahkan podosit dalam jaringan mikro glomerulus (Gambar S7A , Informasi Pendukung). Hal ini menunjukkan bahwa lingkungan 3D, bersama dengan interaksi endotel, memengaruhi ekspresi gen-gen ini. Namun, ekspresi nefrin (NPHS1), molekul pensinyalan dan komponen kunci diafragma celah podosit yang menghubungkan FP, tidak terdeteksi. Hal ini sejalan dengan pengamatan sebelumnya tentang tingkat ekspresi rendah NPHS1 pada ciPOD. [ 18 ] Hal ini mungkin menunjukkan kegagalan podosit untuk membentuk diafragma celah pada model kami. Selain itu, ekspresi faktor transkripsi unik seperti Tbx3 dan Gata5, yang baru-baru ini diidentifikasi sebagai spesifik untuk GEC manusia, [ 19 ] tidak terdeteksi baik pada GEC berlapis 2D maupun pada model 3D kami.
GBM disekresikan dan diremodel oleh podosit dan GEC. Laminin dan kolagen tipe-IV adalah komponen utama GBM dan tidak seperti di sebagian besar membran dasar, isoform mereka berubah selama pematangan glomerulus. [ 20 ] Pada glomerulus dewasa, COL4α1α2α1 terutama digantikan oleh COL4α3α4α5 yang disekresikan oleh podosit, sedangkan LAM1α1β1γ digantikan oleh LAM5α2β1γ, yang disekresikan bersama oleh podosit dan GEC. [ 20 ] Dengan demikian, komposisi GBM dan kontribusi dari GEC dan podosit mencerminkan status kematangan glomerulus. Oleh karena itu, kami mempertanyakan apakah podosit dan GEC merombak lingkungan mikro mereka yang diperkaya kolagen-I secara spesifik dengan mengekspresikan dan mensekresi komponen GBM yang matang. Dalam hal ini, kami memeriksa tingkat ekspresi gen COL4α1 dan LAMA1 sebagai komponen GBM yang belum matang, dan COL4α3 dan LAMA5, yang ditemukan dalam GBM yang matang. Kami mendeteksi ekspresi yang diperkaya untuk semua komponen dalam jaringan mikro glomerulus dibandingkan dengan jaringan mikro GEC dan GEC dan podosit yang dikultur 2D. Ekspresi COL4α3 dan LAMA1 dalam GEC yang dikultur 2D dan jaringan mikro GEC berada di bawah tingkat deteksi, yang menunjukkan bahwa podosit adalah produsen utama LAMA1 dan COL4α3 dalam jaringan mikro glomerulus ( Gambar 3A ) . Di sisi lain, jaringan mikro GEC menunjukkan tingkat ekspresi LAMA5 dan COL4α1 yang lebih rendah daripada jaringan mikro glomerulus namun mengungkapkan kontribusi kedua jenis sel terhadap sintesis. Kami juga menunjukkan bahwa Col-IV disekresikan dan disimpan dalam jaringan mikro sudah 3 hari pasca-penambahan podosit dengan imunofluoresensi. Kami mewarnai mikrotissue dengan antibodi yang mengikat semua isoform Col-IV, dan mikrotissue GEC dan mikrotissue glomerular diwarnai secara positif yang menunjukkan Col-IV ekstraseluler (Gambar 3C,D ). Deposisi laminin tidak diperkaya dalam lingkungan ekstraseluler, berbeda dengan Col-IV (Gambar S7B , Informasi Pendukung).
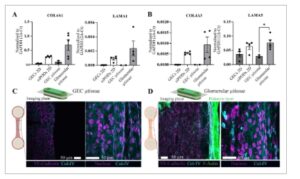
Gambar 3
Buka di penampil gambar
Kekuatan Gambar
Ekspresi dan sintesis komponen GBM dalam mikrotisu 3D. A) Analisis tingkat ekspresi gen untuk COL4A1 dan LAMA1, yang menunjukkan pembentukan GBM yang belum matang. B) Profil ekspresi gen untuk COL4A3 dan LAMA5, yang terkait dengan pematangan GBM. Penilaian imunofluoresensi mengungkapkan keberadaan Col-IV yang kuat di mikrotisu glomerulus C) GEC dan D). Lapisan podosit dengan intensitas F-aktin yang tinggi terlihat jelas di mikrotisu glomerulus. Data dilaporkan sebagai mean ± SD. n = 2 pengulangan independen untuk mikrotisu GEC, dan n ≥ 3 untuk kelompok lain; * P = 0,005, dengan ANOVA satu arah.
2.3 Peregangan dengan Tekanan Tinggi Menyebabkan Terpisahnya Podosit dan Penurunan Kekuatan Kontraktil
Filtrasi glomerulus dipertahankan oleh gradien tekanan hidrostatik. Dengan demikian, dinding kapiler glomerulus terus-menerus diregangkan karena perbedaan antara tekanan hidrostatik glomerulus dan tekanan dalam ruang Bowman ( Gambar 4 A-i ). [ 21 ] Kondisi patologis seperti hipertensi glomerulus atau hiperfiltrasi di mana stres mekanis hemodinamik tinggi berkelanjutan dalam waktu telah terbukti memengaruhi kesehatan podosit dan mendukung penghapusan FP dan pelepasan podosit dari GBM. [ 22 , 23 ] Stres melingkar dan stres aksial dalam kapiler glomerulus yang sehat diketahui masing-masing ≈50 kPa dan 0,3 kPa. [ 24 ]
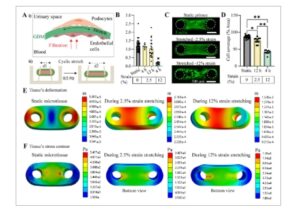
Gambar 4
Buka di penampil gambar
Kekuatan Gambar
Peregangan siklik jaringan mikro glomerulus. Ai) Filtrasi darah memberikan tekanan mekanis pada dinding kapiler dan podosit terpapar tekanan geser tangensial dan tekanan melingkar (peregangan). ii) Ilustrasi yang menunjukkan strategi penerapan peregangan siklik pada jaringan mikro glomerulus dengan frekuensi 0,5 Hz untuk meniru tekanan mekanis yang merupakan komponen penting fisiologi glomerulus. B) Jaringan mikro menunjukkan gaya yang jauh lebih rendah di bawah peregangan mekanis dengan regangan linier yang tinggi (12%). Data dilaporkan sebagai mean ± SD. n ≥ 9 jaringan mikro; * P < 0,001 jika dibandingkan dengan kondisi statis dengan ANOVA satu arah dengan uji Tukey. C) Pencitraan confocal mengungkapkan efek merugikan dari peregangan mekanis dengan regangan linier 12 bahkan setelah 4 jam sedangkan tidak ada efek yang jelas pada organisasi sitoskeletal dan cakupan jaringan mikro yang diamati ketika regangan peregangan adalah 2,5%. Skala batang adalah 500 µm untuk semua gambar. D) Plot yang menunjukkan cakupan yang jauh lebih rendah setelah peregangan mekanis berdasarkan analisis citra fluoresensi. Data dilaporkan sebagai mean ± SD. n ≥ 9 mikrojaringan; * P = 0,001, ** P < 0,0001 jika dibandingkan dengan kondisi statis dengan ANOVA satu arah dengan uji Tukey. E) Model elemen hingga membantu dalam memahami efek deformasi jaringan di bawah peregangan. Deformasi total pada tinggi rata-rata mikrojaringan saat tidak ada regangan, selama peregangan regangan 2,5% dan peregangan regangan 12%. F) Representasi multi-tampilan dari kontur tegangan ekivalen Von Mises yang menunjukkan distribusi tegangan di mana warna merah menunjukkan area dengan tegangan lebih tinggi. Selama peregangan, tegangan sebagian besar terkonsentrasi di sekitar wilayah tempat mikrojaringan menyentuh bagian dalam pilar, yang menunjukkan titik fokus ketegangan mekanis.
Mikrojaringan glomerulus kita terbentuk di sekitar pilar PDMS yang fleksibel, terus-menerus terpapar pada ketegangan pasif karena resistensi mikropilar terhadap pemadatan. Gaya perlawanan setara dengan gaya kolektif yang diberikan oleh sel-sel, yang menarik matriks dan memadatkannya. Kami memperkirakan ketegangan pasif sebagai gaya statis yang dinormalisasi oleh luas penampang mikrojaringan, yang diperoleh dari lebar mikrojaringan dan diasumsikan melingkar. [ 13 ] Kami menemukan nilai ketegangan mikrojaringan sebagai 582 ± 146 Pa (n = 33) (Gambar S8A , Informasi Pendukung). Selain ketegangan pasif ini, kami menyelidiki efek peregangan uniaxial siklik aktif dari mikrojaringan pada gaya seluler yang dihasilkan dan perilaku podosit (Gambar 4A-ii ). Peregangan mekanis dilakukan dengan menempatkan dan memperbaiki perangkat PDMS ke dalam unit tandu, yang terhubung ke pengontrol gerakan, dengan klem (Gambar S8B , Informasi Pendukung). % Regangan pada PDMS diukur untuk gerakan tandu yang berbeda (antara 2 mm-0,5 mm) sebelum peregangan jaringan mikro untuk menilai transmisi regangan target pada posisi yang berbeda (Gambar S8C , Informasi Pendukung). Kami tidak menemukan perbedaan yang signifikan dalam peregangan antara posisi yang berbeda di perangkat (Gambar S8D,E , Informasi Pendukung). Kemudian, kami memaparkan jaringan mikro pada peregangan amplitudo rendah (regangan 2,5%) dan amplitudo tinggi (regangan 12%) untuk berbagai periode dan kami mengukur gaya pada akhir siklus total. Pemaparan jaringan mikro pada peregangan mekanis amplitudo tinggi menyebabkan penurunan gaya yang signifikan (penurunan menjadi 0,16 dari nilai sebelum peregangan) sedangkan peregangan regangan rendah tidak menyebabkan perubahan gaya yang signifikan (Gambar 4B ). Ketika kami menyelidiki secara dekat jaringan mikro dengan mikroskopi confocal, pelepasan podosit dari jaringan mikro terbukti dalam kelompok peregangan amplitudo tinggi dibandingkan dengan peregangan amplitudo rendah dan kondisi kontrol statis (Gambar 4C ; Gambar S9 , Informasi Pendukung). Kuantifikasi cakupan podosit dengan analisis gambar juga mengonfirmasi pengurangan signifikan pada area yang ditutupi sel dalam peregangan amplitudo tinggi dengan area tertutup sebesar % 41,3 ± 6,0 dibandingkan dengan % 75,3 ± 8,6 dan % 86,7 ± 5,1 untuk peregangan regangan 2,5% dan kontrol statis, masing-masing (Gambar 4D ).
Kami juga telah mensimulasikan deformasi dan distribusi tegangan untuk jaringan mikro statis dan ketika jaringan mikro diregangkan pada frekuensi yang berbeda dengan analisis mekanis terbatas (Gambar S10 , Informasi Pendukung). Simulasi ini mengungkapkan pola tegangan di seluruh jaringan mikro ketika mereka mengalami tingkat peregangan yang berbeda. Kami menggabungkan tinggi rata-rata jaringan mikro, seperti yang diamati dalam percobaan kami, ke dalam analisis untuk meningkatkan akurasi simulasi. Hasilnya menunjukkan bahwa deformasi jaringan yang paling signifikan terjadi di daerah tengah jaringan mikro, sedangkan dalam kondisi statis, deformasi lebih besar di daerah luar di sekitar pilar mikro (Gambar 4E ). Selain itu, simulasi tegangan yang diberikan selama peregangan mengungkapkan tingkat tegangan yang lebih tinggi di daerah sekitar pilar mikro, dengan nilai puncak mencapai 128 kPa untuk kelompok peregangan 12%. Nilai-nilai ini melebihi rata-rata yang sehat dan menyerupai kondisi hipertensi. Sebaliknya, peregangan 2,5% dan kondisi statis menghasilkan tegangan tarik ≈20 kPa dan 0,3 kPa, dengan nilai puncak masing-masing 34 dan 0,5 kPa, (Gambar 4F ) yang selaras dengan tegangan darah normal. Hasil eksperimen untuk kelompok peregangan 12% menunjukkan pelepasan sel dan berkurangnya cakupan jaringan mikro di area yang sama, yang menunjukkan bahwa integritas struktural terganggu saat mengalami tegangan berulang tingkat tinggi (Gambar S9 , Informasi Pendukung). Secara keseluruhan, hasil ini menyoroti potensi efek merugikan dari beban mekanis tinggi pada ciPod kami. Lebih jauh lagi, peregangan mekanis dapat digunakan untuk meniru kerusakan akibat hipertensi dan patofisiologi untuk penelitian di masa mendatang.
2.4 Pengobatan Podosit dengan Sitokin TNFα yang Relevan terhadap Penyakit Menyebabkan Penurunan Kekuatan Mekanik pada Mikrojaringan
Tumor necrosis factor alpha (TNFα) telah terlibat dalam perkembangan dan progresi glomerulosklerosis fokal dan segmental (FSGS), suatu bentuk penyakit ginjal kronis (CKD) yang parah yang ditandai dengan jaringan parut pada glomerulus. Jalur TNFα intrinsik podosit telah terbukti diaktifkan pada sebagian kecil pasien FSGS dan dikaitkan dengan hasil ginjal yang buruk. [ 25 , 26 ] In vitro, podosit yang terpapar plasma FSGS menunjukkan peningkatan ekspresi gen jalur TNFα, gangguan sitoskeleton, dan aktivasi integrin β3, dan efek ini dibalikkan oleh blokade TNFα. [ 27 ] Selain itu, TNFα menginduksi ekspresi gen pro-apoptosis RARRES1 pada podosit yang dikultur, yang ekspresinya berkorelasi positif dengan penurunan fungsi ginjal pada penyakit glomerulus manusia. [ 28 ] TNFα dan reseptornya, khususnya TNFR2, yang diekspresikan dalam podosit manusia dan GEC, telah diusulkan sebagai biomarker untuk memprediksi hasil ginjal pada FSGS. Kadar TNFR2 yang meningkat berkorelasi dengan penurunan fungsi ginjal dan dapat berfungsi sebagai indikator awal perkembangan CKD. [ 29 ] Oleh karena itu, kami menguji TNFα sebagai sitokin yang relevan dengan penyakit untuk podositopati dan menyelidiki bagaimana paparan TNFα memengaruhi aktivitas remodeling sitoskeletal podosit dengan mengukur perubahan pada kekuatan kontraktil podosit. Pertama, kami melakukan uji titrasi dosis untuk menentukan konsentrasi TNFα optimum yang menginduksi kerusakan podosit tanpa menyebabkan toksisitas kritis dan menginduksi kerusakan sitoskeletal yang dapat dinilai dengan pembacaan gaya (Gambar S11 , Informasi Pendukung). Akibatnya, pengobatan TNFα 200 ng ml −1 dimulai 1 hari setelah penambahan podosit dan dipertahankan selama 7 hari ( Gambar 5 A ). Kami awalnya mengamati peningkatan gaya ketika masing-masing mikrotissu dinormalisasi ke nilai sebelum perawatan, dan pengurangan gaya dimulai setelah 5 hari paparan TNFα (Gambar 5B ). Kami menemukan bahwa podosit masih menunjukkan ekspresi vinculin yang terlokalisasi, meskipun pada tingkat yang berkurang yang membuktikan bahwa mereka tetap melekat pada mikrotissu (Gambar S12 , Informasi Pendukung). Ini mungkin menunjukkan bahwa transmisi gaya seluler dan perlekatan sel adalah fenotipe yang dapat dipisahkan, mirip dengan yang dilaporkan setelah model perawatan aminonukleosida puromisin untuk sistem 2D. [ 30 ] Selain itu, paparan TNFα menghasilkan morfologi sel yang lebih memanjang dan terpolarisasi (Gambar 5C ; Gambar S13), Informasi Pendukung). Perubahan fenotipik ini dikaitkan dengan penurunan kekuatan kontraktil yang dihasilkan oleh podosit. Secara keseluruhan, perubahan ini menunjukkan potensi hilangnya ketahanan biomekanik mikrojaringan setelah pengobatan TNFα sambil mempertahankan viabilitas podosit. Namun, ketika mikrojaringan diobati dengan 0,1% Triton X-100, yang berfungsi sebagai kontrol positif untuk kerusakan podosit dan viabilitas yang terganggu, penurunan kekuatan yang signifikan, bersama dengan pelepasan sel dan morfologi yang terganggu, diamati (Gambar S14 , Informasi Pendukung).
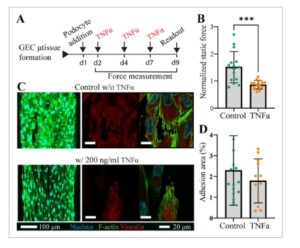
Gambar 5
Buka di penampil gambar
Kekuatan Gambar
Cedera podosit yang disebabkan oleh TNFα. A) Tinjauan umum strategi untuk memaparkan jaringan mikro pada 200 ng ml −1 TNFα selama 7 hari. B) Plot gaya jaringan mikro statis yang diukur pada hari ke-9 dinormalisasi dengan pengobatan pra-TNFα. Lipatan tersebut menunjukkan pengurangan gaya yang disebabkan oleh TNFα. C) Gambaran imunofluoresensi representatif dari nuklei, aktin, dan vinculin pada hari ke-9, dengan atau tanpa pengobatan TNFα. Morfologi memanjang yang diinduksi pada podosit (panel kiri). Pewarnaan vinculin juga menunjukkan intensitas yang lebih rendah dibandingkan dengan jaringan mikro kontrol ketika jaringan mikro diobati dengan TNFα. D) Plot area adhesi (%) yang dihitung berdasarkan pewarnaan vinculin menunjukkan penurunan pada kelompok kerusakan yang disebabkan oleh TNFα. Data dilaporkan sebagai mean ± SD. n ≥ 7 jaringan mikro/pengulangan; diulang setidaknya 3 kali. Nilai P dihitung dengan uji-t, ***P < 0,001.
2.5 Ekspresi Berlebih TRPC6 Menggantikan Mutasi TRPC6 yang Berfungsi sebagai Target Terapi dan Menyebabkan Pengurangan Kekuatan Mekanik
Pengaturan kalsium intraseluler sangat penting untuk fungsi fisiologis podosit karena masuknya kalsium yang berlebihan dapat mengakibatkan penghapusan prosesus kaki, apoptosis, dan kerusakan. [ 7 , 8 , 31 ] Mutasi perolehan fungsi TRPC6 manusia telah dilaporkan menyebabkan masuknya kalsium yang berlebihan pada podosit, memainkan peran penting dalam patogenesis penyakit ginjal. [ 8 ] Disregulasi dalam masuknya kalsium tersebut memengaruhi dinamika aktin, yang mengakibatkan de-arrangemen sitoskeletal, distribusi F-aktin yang abnormal, dan cedera podosit. [ 7 ] Untuk mengonfirmasi relevansi model kami dan mengeksplorasi potensi jaringan mikro kami sebagai platform validasi target, kami menguji podosit immortal dengan ekspresi berlebih TRPC6 yang diinduksi Dox untuk mensimulasikan peningkatan masuknya kalsium yang diamati dengan mutasi perolehan fungsi TRPC6 ( Gambar 6 A-i ; Gambar S15 , Informasi Pendukung). Mikrojaringan glomerulus yang terbentuk dengan garis podosit oeTRPC6 menunjukkan morfologi yang mirip dengan mikrojaringan tipe liar (Gambar 6A-ii ). Analisis mikroskopi imunofluoresensi konfokal mengonfirmasi ekspresi dan keberadaan TRPC6 dalam mikrojaringan yang diinduksi Dox (Gambar 6A-ii ). Selain itu, mikrojaringan yang diinduksi Dox menunjukkan ekspresi gen TRPC6 yang sangat tinggi, sedangkan tingkat ekspresi dalam mikrojaringan yang tidak diinduksi tidak terdeteksi (Gambar S15 , Informasi Pendukung).
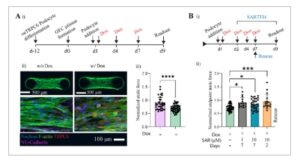
Gambar 6
Buka di penampil gambar
Kekuatan Gambar
Rekapitulasi cedera podosit yang diinduksi secara genetik yang disebabkan oleh TRPC6. Ai) Tinjauan umum strategi dan garis waktu untuk menginduksi cedera yang disebabkan oleh TRPC6. ii) Gambaran konfokal yang membandingkan jaringan mikro yang terbentuk tanpa atau dengan adanya Dox untuk menginduksi ekspresi berlebihan TRPC6, yang tidak berpengaruh pada morfologi jaringan mikro dan cakupan oleh podosit. Induksi Dox menghasilkan ekspresi TRPC6 yang kuat (merah) dibandingkan dengan kelompok tanpa induksi seperti yang diilustrasikan oleh gambar fluoresensi yang representatif. iii) Plot yang menunjukkan distribusi gaya kontraktil (dinormalisasi hingga 1 hari pasca penyemaian podosit) dan penurunan yang signifikan ketika TRPC6 diinduksi dan diekspresikan secara berlebihan. Nilai P dihitung dengan uji-t dua sisi, n ≥ 20 jaringan mikro, **** P < 0,001. Bi) Tinjauan umum strategi penanganan senyawa penghambat untuk skema pencegahan dan penyelamatan, ii) plot yang menunjukkan distribusi gaya kontraksi untuk jaringan mikro yang diobati dengan SAR7334 1µM dan 10 µM dan tanpa pengobatan. Penanganan SAR7334 menghambat pengurangan gaya yang disebabkan oleh ekspresi TRPC6 yang berlebihan. Nilai P dihitung dengan ANOVA satu arah, n ≥ 6 jaringan mikro per pengulangan, *** P < 0,001. Eksperimen penanganan SAR diulang 2 kali.
Ketika kami menganalisis kekuatan kontraktil, tidak ada perbedaan yang jelas 3 hari pasca-induksi; namun, jaringan mikro glomerulus menunjukkan penurunan yang signifikan dalam kekuatan kontraktil (penurunan menjadi 0,66 dari nilai 1 hari pasca-induksi) pada 8 hari pasca-induksi ketika diinduksi dengan Dox dibandingkan dengan jaringan mikro yang tidak diinduksi (menurun menjadi 0,98) (Gambar 6A-iii ). Kami juga menyelidiki apakah penambahan senyawa dalam rejimen pengobatan pencegahan (1 hari pasca-induksi Dox) memengaruhi organisasi F-actin dalam jaringan mikro glomerulus 3D dan kultur podosit 2D. Dalam kedua kasus, setiap perubahan yang nyata tidak diamati dalam hal struktur sitoskeleton dengan penambahan Dox atau dengan pengobatan SAR7334 (Gambar S16 , Informasi Pendukung).
Selain itu, perubahan dalam lebar jaringan tidak menunjukkan perbedaan yang signifikan dan jaringan mikro yang diinduksi Dox kurang padat dibandingkan dengan yang tidak diinduksi. Temuan ini selaras dengan laporan saat ini, karena ekspresi berlebihan TRPC6 menyebabkan pengurangan kekuatan kontraktil podosit, berkorelasi dengan sitoskeleton aktin yang tidak stabil. Mengingat validitas pembacaan gaya, strategi kami memungkinkan rekapitulasi cedera yang diinduksi secara kimia dan genetik dalam lingkungan mikro 3D, untuk validasi target lebih lanjut dari target obat baru dan pengujian senyawa potensial. Dalam hal ini, untuk menyelidiki perlindungan dan penyelamatan gaya yang diberikan oleh podosit, kami merawat jaringan mikro dengan penghambat saluran TRPC6 yang diketahui, SAR7334. Dalam rejimen pengobatan pencegahan, pengobatan penghambat TRPC6 dimulai 1 hari pasca induksi Dox dan dipertahankan selama periode kultur (Gambar 6B-i ). Perlakuan SAR7334 pada 1 dan 10 µM secara kuat mencegah penurunan gaya pada jaringan mikro dan mempertahankan tingkat gaya jaringan yang serupa dengan kelompok kontrol tanpa perlakuan yang tidak diinduksi, di mana TRPC6 tidak diekspresikan (Gambar 6A-iii,B-ii ). Untuk model terapeutik, kami membiarkan cedera berkembang selama 5 hari dengan induksi Dox sebelum pemberian senyawa penghambat. Kami kemudian merawat jaringan mikro dengan SAR7334 10 µM selama 2 hari sambil mempertahankan perlakuan Dox. Mirip dengan skema perlakuan pencegahan, kami mengamati penyelamatan gaya bahkan setelah 2 hari pada kelompok perlakuan dibandingkan dengan kontrol tanpa perlakuan. Hasilnya, gaya dipertahankan sekitar nilai penyemaian podosit 1 hari pasca (perubahan 1,1 kali lipat). Bersama-sama, data ini menunjukkan efek perlakuan SAR7334 dalam menghambat aktivitas berlebihan TRPC6 dan destabilisasi aktin yang dihasilkan, sehingga menyelamatkan gaya yang dihasilkan oleh aktin sitoskeleton. Model pengobatan dan cedera genetik ini menunjukkan penerapan dan potensi jaringan mikro glomerulus kami untuk digunakan dalam pengujian senyawa untuk target yang memengaruhi stabilitas dan organisasi sitoskeletal. SAR7334 dipilih untuk demonstrasi pembuktian konsep ini karena potensi nanomolar dan bioavailabilitasnya. [ 32 ] Model ini juga dapat digunakan untuk mengevaluasi inhibitor TRPC6 dan senyawa baru lainnya yang diketahui, menyediakan alat serbaguna untuk pengujian senyawa.
3 Diskusi
Ginjal, dengan jenis selnya yang beragam, mekanika filtrasi yang rumit, dan organisasi spasial yang tepat, menyajikan kompleksitas anatomi yang menantang untuk ditiru dalam model. Meskipun kemajuan terbaru dalam model organoid ginjal dan glomerulus spheroid telah berhasil mereplikasi interaksi sel-ke-sel dan mencapai kematangan podosit yang ditingkatkan, model-model ini sering kali tidak memiliki GBM yang matang serta integrasi isyarat biomekanik aktif yang diperlukan untuk mempelajari ketahanan biomekanik podosit. [ 9 , 18 ] Model in vitro rekayasa hayati 3D dari glomerulus manusia, yang menggabungkan GBM matang yang disekresikan sel atau dideselularisasi, memfasilitasi studi penyakit ginjal kronis terkait GBM. [ 33 , 34 ] Namun, model-model ini masih kekurangan investigasi ke dalam ketahanan biomekanik dinamis podosit. Akhirnya, sistem mikrofisiologis, menyediakan perfusi dan mencoba mereplikasi mekanobiologi glomerulus tetapi sering kali tidak menggabungkan kelengkungan podosit dan pengukuran gaya kontraktil, yang penting untuk pemahaman yang komprehensif tentang fungsi ginjal. Mengembangkan model in vitro yang mereplikasi mekanobiologi podosit dan mengevaluasi hilangnya ketahanan biomekanik pada penyakit sangat penting untuk mengidentifikasi target dan menciptakan terapi presisi untuk podositopati. Hingga saat ini, susunan mikrotissu 3D telah diusulkan untuk mengembangkan jaringan otot, mempelajari mekanobiologi dan remodeling matriks, dan pengujian obat anti-fibrotik. [ 13 , 35 – 38 ] Di sini, kami menyajikan susunan mikrotissu 3D manusia canggih yang dirancang untuk merangkum struktur tiga lapis glomerulus termasuk tegangan tarik pada podosit. Kami menunjukkan bahwa menempatkan podosit ke dalam lingkungan 3D meningkatkan ekspresi gen terkait podosit mereka mungkin karena efek lingkungan mikro seperti crosstalk dengan sel endotel dan tegangan mekanis yang ada di antara mikropilar. Telah ditunjukkan bahwa podosit bersifat mekanosensitif dan merespons peregangan mekanis dengan menghasilkan tonjolan sel yang panjang dan tipis, mengatur ulang aktin cytoskeleton, dan melepaskan diri dari membran dasar. [ 39 – 41 ] Peregangan jaringan mikro, yang meniru tekanan melingkar pada dinding kapiler, menyebabkan pelepasan podosit yang signifikan dan hilangnya ketegangan dari jaringan mikro tersebut, yang sejalan dengan laporan terkini tentang mekanobiologi podosit 2D. Pemeriksaan proses ini dalam jaringan mikro 3D pada berbagai jangka waktu dan tingkat ketegangan dapat memberikan wawasan berharga tentang aspek biofisik patologi glomerulus dan menjelaskan studi tentang terapi baru.
Platform mikrojaringan ini dengan desain mikropilarnya yang fleksibel memungkinkan pengukuran gaya seluler secara langsung sebagai indikator fungsi sitoskeletal podosit dan kesehatan secara keseluruhan, yang terganggu dalam berbagai podositopati. Selain itu, platform ini memungkinkan pemodelan cedera podosit yang disebabkan oleh bahan kimia dan genetik dan berfungsi sebagai alat untuk validasi target obat baru di masa mendatang dan untuk memprediksi kemanjuran pengobatan senyawa yang memodulasi efek target potensial. Dengan demikian, kami menguji penerapan sistem ini untuk mengevaluasi efek target dengan hubungan genetik dengan podositopati seperti TRPC6 dan menunjukkan untuk pertama kalinya secara in vitro penurunan gaya kontraktil yang diberikan oleh podosit yang mengekspresikan TRPC6 secara berlebihan. Selain itu, kami menunjukkan kemanjuran terapeutik senyawa penghambat TRPC6, SAR7334, yang mencegah pengurangan gaya dalam model cedera.
Hasil ini menunjukkan bahwa jaringan mikro glomerulus menghadirkan strategi yang ampuh untuk merangkum cedera podosit yang disebabkan oleh mekanik, kimia, dan genetik dan dapat memberikan wawasan tentang opsi pengobatan yang potensial. Platform jaringan mikro kami, bersama dengan titik akhir pengganti yang relevan dengan penyakit (kekuatan kontraksi podosit dan pelepasan podosit dari GBM) juga menjanjikan untuk digunakan guna mengatasi berbagai jenis pertanyaan mekanobiologis dalam podosit yang tidak dapat diatasi oleh metode kultur sebelumnya. Dalam hal ini, sinergi antara teknologi organoid dan platform jaringan mikro dapat dibangun dan sel-sel yang berasal dari organoid pasien, bukan lini sel yang diabadikan, dapat diimplementasikan untuk menawarkan model praklinis yang lebih kuat. Selain itu, model jaringan mikro dapat dengan mudah diadaptasi untuk hasil yang lebih tinggi untuk aplikasi penyaringan obat dengan pembacaan mikroskopisnya yang mudah. Pengulangan desain di masa mendatang juga dapat memungkinkan pembuatan jaringan mikro dengan bentuk dan organisasi yang lebih kompleks untuk jaringan yang berbeda. [ 42 ]
Baru-baru ini, ditunjukkan bahwa integrasi aliran meningkatkan vaskularisasi organoid ginjal dan pematangan podosit dan kompartemen tubulus di dalamnya. [ 43 ] Mengembangkan sistem perfusi sangat penting untuk meniru keadaan fisiologis dan patofisiologis glomerulus ginjal dengan lebih baik. Model jaringan mikro kami tidak perfusi; oleh karena itu, ia tidak memiliki tegangan geser yang biasanya dialami oleh sel endotel dan podosit. Memperkenalkan tegangan geser dan perfusi ke dalam sistem jaringan mikro juga dapat memberikan kesempatan untuk mengkorelasikan hilangnya gaya dengan peningkatan kebocoran albumin ke dalam urin. Pada tingkat teknis, batasan lain dari model kami saat ini, karena organisasi spasial sel endotel dalam jaringan mikro, adalah kurangnya penilaian fungsi penghalang, khususnya yang berkaitan dengan permeabilitas albumin. Juga, dalam model ini, ciPOD digunakan sebagai sumber podosit; namun, mereka tidak memiliki ekspresi protein diafragma celah. Oleh karena itu, kami mengukur gaya yang diberikan oleh jaringan aktin kontraktil di dalam badan sel, tetapi kami tidak dapat memahami kontribusi dari elemen nonkontraktil yang ada dalam prosesus kaki terhadap ketahanan biomekanik podosit. Penerapan garis podosit yang menampilkan FP dan mengekspresikan protein diafragma celah, alih-alih ciPOD, dalam penelitian mendatang dapat memungkinkan kami untuk memahami kontribusi dan hubungan elemen nonkontraktil terhadap gaya yang diberikan untuk menyeimbangkan peregangan hemodinamik.
4 Bagian Eksperimen
Kultur Sel dan Pemeliharaan
Podosit manusia yang diabadikan secara bersyarat (ciPOD) dari Dr. Moin Saleem dari Universitas Bristol (1) dikultur pada suhu permisif 33 °C dalam 5% CO2 dengan menggunakan medium lengkap dengan RPMI1640 yang mengandung L-glutamin dan Natrium Bikarbonat (Sigma-Aldrich) yang dilengkapi dengan 1% insulin transferrin selenium (Thermo Fisher), 1% Penisilin/Streptomisin (GIBCO) dan 10% serum sapi janin (Sigma). Sel-sel ditransportasikan dua kali seminggu dengan pengobatan dengan 0,05% tripsin-EDTA (GIBCO). Sel Endotel Mikrovaskular Glomerulus Primer Manusia (ACBRI 128, Sistem Sel) dikulturkan pada pelat berlapis kolagen tipe 1 (PureCol, Advanced Biomatrix) menggunakan medium lengkap Endothelial Cell Growth Medium yang dilengkapi dengan MV 2 KIT (Promocell C-22121) pada suhu 37 °C dalam 5% CO 2 . Cawan kultur jaringan dilapisi terlebih dahulu dengan 50 µg ml −1 PureCol dalam larutan penyangga fosfat (PBS)-/- selama 20 menit pada suhu 37 °C.
Sel Endotel Vena Umbilika Manusia (HUVEC) dikultur dalam Medium Pertumbuhan Sel Endotel yang dilengkapi dengan MV 2 KIT pada suhu 37 °C dalam 5% CO 2 .
Pelapisan Laminin dan Induksi Diferensiasi ciPOD
Untuk induksi fenotipe podosit, ciPOD disemai pada pelat berlapis Laminin. Labu kultur jaringan T75 dilapisi dengan Laminin521 (Biolamina MSPPLN-521-05) pada pengenceran 1:40 dalam PBS+/+ selama 1,30 jam pada suhu 37 °C. Sel kemudian disemai pada kepadatan 1,2 M per T75 dan dikultur pada suhu 37 °C dalam 5% CO2 selama minimal 10 hari dengan penggantian media 2–3 kali per minggu.
Persiapan Perangkat PDMS dan Pembentukan Mikrojaringan
Perangkat PDMS yang dirancang khusus berisi 18 sumur kecil dengan dimensi 3,6×1,8×2 mm (pxlxt), masing-masing dengan dua pilar fleksibel (diameter 500 µm, tinggi 2,5 mm, jarak pusat-ke-pusat 1,8 mm) (Gambar S1 , Informasi Pendukung), dibeli dari IBIDI (Jerman).
Sebelum gel dan penyemaian sel, ruang PDMS disterilkan di bawah sinar UV selama 1 jam dan kemudian dilapisi semalam dengan 2% v/v Pluronic F127 (Sigma-Aldrich) dalam air untuk mencegah adhesi protein matriks ke dinding PDMS. Setelah menyedot larutan, 6 µl larutan kolagen-I dingin yang tidak dipolimerisasi dipipet ke dalam setiap sumur. Larutan 100 µl berisi 20 µl Kolagen I 5 mg ml −1 (Cultrex, R&D Systems), 10 µl HEPES 1 M (Gibco), 60 µl media kultur, dan 10 µl NaHCO 3 (37 g l −1 , BioConcept). Perangkat PDMS disentrifugasi (1 menit pada 1000 rpm) dan diinkubasi pada suhu kamar selama 30 menit untuk memungkinkan polimerisasi. Berikutnya, lapisan kedua 6 µl larutan kolagen-I dingin tak terpolimerisasi yang dicampur dengan 30000 GEC dipipet di atas lapisan bawah terpolimerisasi di setiap ruang PDMS dan dibiarkan berpolimerisasi selama 30 menit pada 37 °C dan 5% CO 2 . Kemudian, 1 ml media kultur ditambahkan ke setiap perangkat untuk memungkinkan pembentukan jaringan mikro semalaman. ciPOD pada kepadatan 4×10^5 sel ml −1 ditambahkan di atas jaringan mikro GEC yang sudah terbentuk setelah 24 jam, dan jaringan mikro disimpan dalam media kultur khusus GEC. 2 µM CellTrace Calcein Green (Invitrogen) digunakan untuk memberi label ciPOD sebelum penyemaian untuk menilai perlekatannya pada jaringan mikro.
Pengukuran Kekuatan Mikrojaringan
Lendutan pilar δ digunakan untuk mengukur gaya kontraktil aksial F dari setiap jaringan mikro menggunakan hukum Hooke F = k × δ, di mana k adalah konstanta pegas pilar PDMS pada ketinggian jaringan di atas dasar pilar h jaringan . k diberikan oleh produsen, dan diukur pada ketinggian pilar yang berbeda dan ditentukan dari hubungan gaya-tekukan.
Untuk menghitung gaya berdasarkan lendutan pada ketinggian yang berbeda (h), persamaan balok Euler-Bernoulli F h = k 0 × δ/( h / h 0 ) 3 di mana h 0 adalah tinggi acuan yang ditetapkan sebesar 1 mm, dan k 0 adalah konstanta pegas pilar pada ketinggian titik acuan. Konstanta pegas k adalah 11 µN µm −1 .
Agar lebih akurat, defleksi jaringan mikro di bagian atas pilar dan defleksi terhitung δ di tingkat jaringan diukur menggunakan persamaan berikut.δ = δ top × h jaringan /(3 × h top − h jaringan ), di mana h top adalah tinggi pilar.
Peregangan Jaringan Mikro
Peregangan jaringan mikro dilakukan sebagai pengganti stres yang dihasilkan dalam penghalang filtrasi glomerulus karena filtrasi cairan dan tekanan hidrostatik dalam kapiler. Untuk menerapkan peregangan dalam rentang fisiologis, perangkat PDMS dimasukkan ke dalam sistem peregangan yang dibuat sendiri (Gambar S7 , Informasi Pendukung) yang dihubungkan ke pengontrol gerakan yang dapat diprogram (C-867, Physik Instrumente (PI) GmbH, Jerman). Regangan siklik diterapkan pada jaringan mikro pada frekuensi 0,5 Hz dan amplitudo peregangan 0,5 mm yang sesuai dengan regangan 2,5 ± 0,5%. Gambar brightfield jaringan mikro sebelum dan sesudah peregangan diperoleh untuk menghitung perubahan gaya yang diberikan oleh podosit.
Analisis Elemen Hingga Deformasi Mikrojaringan
Model 3D dari mikrotissue dan substrat PDMS dibuat oleh SOLIDWORKS 2022 (Dassault Systemes, Prancis), kemudian diimpor ke ANSYS Workbench 2023 R1 untuk mensimulasikan kondisi tegangan. Berdasarkan persyaratan akurasi hasil simulasi dan keseimbangan komputasi, penggabungan multiwilayah akhirnya diterapkan pada model. Matriks seluler dan jaringan sitoskeletal dianggap sebagai material yang sangat viskoelastis, oleh karena itu model neo-Hookean hiperelastis mengacu pada representasi deformasi mikrotissue fleksibel di bawah kontraksi aktif dan peregangan pasif. Parameter elastis isotropik material ditetapkan untuk komponen PDMS. Parameter seperti tinggi jaringan dan gaya kontraktil untuk defleksi jaringan diekstraksi dari eksperimen dan dimasukkan dalam pengaturan simulasi. Gaya kontraksi yang dihasilkan oleh mikrotissue diberikan dalam bentuk beban dukung yang setara di sekitar pilar, di mana kontak terjadi antara permukaan silinder atau di tepinya, dan tekanan yang tidak seragam mungkin ada. Hasil akhir meliputi kontur yang mencerminkan deformasi total, serta kontur distribusi deformasi dan tegangan setiap komponen.
Imunofluoresensi dan Mikroskopi
Jaringan mikro difiksasi dengan 4% paraformaldehida (Electron Microscopy Sciences) dalam perangkat PDMS, dicuci secara serial dengan PBS, dan dipermeabilisasi oleh 0,5% Triton x-100 (Aldrich 23472–9). Pemblokiran kemudian dilakukan dalam 3% BSA (Fluka 05 479) dan jaringan mikro kemudian diinkubasi semalam dengan antibodi primer terhadap VE-Cadherin (BD, 5 61 567, 1:100), TRPC6 (Alomone ACC-017, 1:400), Vinculin (Sigma V9131, 1:200), αvβ3 (Chemicon, MAB1976, 1:200), panci Kolagen IV (Abcam 6585, 1:100) dan diberi label dengan antibodi anti-IgG yang terkonjugasi fluorofor (Thermo Fisher, AlexaFluor, 1:500). Untuk mewarnai sitoskeleton, digunakan probe F-aktin, AlexaFluor 488 Phalloidin dan AlexaFluor 647 Phalloidin (Thermo Fisher) pada pengenceran 1:300. Inti sel diwarnai dengan Hoechst 33 342 (Thermo Fisher, 1:1500).
Mikrojaringan yang diwarnai imun divisualisasikan melalui mikroskop konfokal (FV3000, Olympus, Tokyo), dan gambar diproses menggunakan perangkat lunak analisis gambar Olympus CellSense.
Untuk memvisualisasikan kolagen-I bebas label, digunakan mikroskopi multifoton (Mikroskop Olympus FVMPE-RS, Tokyo) yang dilengkapi dengan lensa objektif 25x. Sampel dieksitasi oleh laser inframerah yang dapat disetel dan tumpukan gambar diperoleh menggunakan generasi harmonik kedua.
Cedera yang Disebabkan oleh TNFα
Cedera yang dimediasi TNFα diinduksi dengan mengobati jaringan mikro dengan 40 dan 200 ng ml −1 TNFα manusia rekombinan (Peprotech 300–01A) yang dilengkapi dengan medium GEC lengkap 1 hari setelah penambahan ciPOD. Kerusakan yang diinduksi TNFα kemudian dibiarkan berkembang selama 7 hari. Medium tanpa TNFα digunakan sebagai kontrol. Medium kultur diganti setiap 2 hari. Setelah 7 hari pengobatan, cedera dinilai menggunakan pembacaan gaya, qPCR waktu nyata, dan imunofluoresensi.
Pembuatan Podosit yang Mengekspresikan TRPC6 Secara Berlebihan dan Pengujian Senyawa
ciPOD dengan TRPC6 yang dapat diinduksi dihasilkan secara in-house melalui transduksi lentivirus menggunakan vektor dengan resistensi blastisidin. Secara singkat, ciPOD disemai pada pelat berlapis laminin, dan 7 jam setelah penyemaian, ciPOD tersebut terinfeksi dengan lentivirus. Seleksi dimulai 2 hari setelahnya dengan Blastisidin pada konsentrasi 10µg mL −1 . Transduksi TRPC6 kemudian dikonfirmasi melalui qPCR. Untuk percobaan mikrojaringan berikut, ciPOD dikultur dan dibedakan seperti yang dijelaskan sebelumnya di bagian kultur sel dan pemeliharaan, kemudian diobati dengan 200 ng ml −1 Doxycycline (Sigma D9891) 24 jam sebelum percobaan untuk menginduksi ekspresi berlebih TRPC6.
Penghambat saluran TRP, SAR7334 (Selleckchem), pengobatan dimulai 1 hari pasca induksi Dox dan dipertahankan sepanjang periode kultur (Gambar 6B-i ). Pengobatan SAR7334 pada 1 dan 10 µM.
Isolasi RNA dan PCR Real-Time Kuantitatif
Isolasi RNA dari jaringan mikro dilakukan dengan menggunakan RNeasy-Micro-Kit (Qiagen, California, AS) sesuai dengan petunjuk pabrik pembuatnya. Sintesis cDNA kemudian dilakukan dengan High-Capacity cDNA Reverse Transcription Kit (Applied Biosystems) dan qPCR dilakukan dengan TaqMan Gene expression Master Mix (Thermo-Fischer) menggunakan QuantStudio 7 Flex Real-Time PCR System (Thermo-Fischer). β-actin dan/atau GAPDH digunakan sebagai gen housekeeping. qPCR dilakukan dengan menggunakan primer TaqMan yang tercantum dalam berkas Informasi Pendukung.
Analisis Statistik
Analisis statistik dilakukan dengan perangkat lunak GraphPad Prism. Data disajikan sebagai rata-rata dengan batang kesalahan yang menunjukkan deviasi standar (SD) kecuali disebutkan sebaliknya (n didefinisikan dalam keterangan setiap gambar). Data dianalisis dengan ANOVA satu arah. Perbedaan signifikansi diverifikasi dengan metode Tukey post-hoc dengan nilai p kurang dari 0,05 kecuali dinyatakan sebaliknya dalam keterangan gambar.